Convergent evolution of berberine biosynthesis
小檗碱生物合成的趋同进化

摘要
小檗碱是一种有效的抗菌和抗糖尿病生物碱,主要从不同植物谱系中提取,特别是从小檗属(毛茛目,早期分支的真双子叶植物)和黄柏属(无患子目,核心真双子叶植物)中提取。与小檗属物种中已知的小檗碱生物合成路径相比,其在黄柏属物种中的生物合成仍不清楚。通过使用染色体水平基因组组装、共表达矩阵和生化测定,我们鉴定了黄柏属小檗碱生物合成中的六个关键步骤,包括甲基化、羟基化和小檗碱桥形成。特别地,我们发现了一类特定的O-甲基转移酶(NOMT),负责N-甲基化。对PaNOMT9的结构分析和诱变研究揭示了其独特的底物结合构象。此外,与毛茛目经典的FAD依赖性小檗碱桥形成不同,黄柏属使用NAD(P)H依赖性的单加氧酶(PaCYP71BG29)进行小檗碱桥的形成,该过程源于色氨酸胺5-羟化酶的新功能化。综上所述,这些发现揭示了小檗碱生物合成在小檗属和黄柏属之间的趋同进化,并标志着趋同进化在植物特化代谢中的作用。
引言
植物来源的苄基异喹啉生物碱(BIA)是药物的一个重要来源。许多BIA,如小檗碱(1–4)、吗啡(5, 6)和血根碱(7, 8),已经用于治疗各种疾病。关于BIA的分离和植物界中的分布,已有大量研究完成。一方面,令人感兴趣的是,许多BIA主要集中在特定的被子植物目中,例如早期分支的真双子叶植物(如毛茛目和木兰目)和木兰类植物(如木兰目、月桂目和胡椒目)(9–12)。另一方面,少数BIA(如小檗碱)的分布则呈现不同的模式。小檗碱最初是从核心真双子叶植物(无患子目)的黄木(Zanthoxylon clava)树皮中分离出来的。后续的研究表明,黄柏属(Phellodendron spp.)也能产生小檗碱(13)。截至目前,小檗碱的生产主要依赖于如毛茛目(Ranunculales)中的小檗属(Berberis)和小檗属(Coptis)以及无患子目(Sapindales)中的黄柏属等植物来源(14)。
小檗碱是一种有效的广谱抗菌和抗糖尿病药物。过去的研究已经描述了小檗碱在小檗属植物根部和根茎中的主要生物合成步骤(15)。其生物合成途径始于多巴胺(1)和4-羟基苯乙醛(4HPAA;2)的偶联反应,生成(S)-去氢可克劳林(3),这一过程由去氢可克劳林合酶(NCS)催化,NCS是PR10/Bet v1家族的成员(16, 17)。随后的修饰反应,包括通过O-/N-甲基转移酶(MTs)进行的O/N-甲基化和通过细胞色素P450进行的羟化,促使(S)-网状碱(7)的形成(18–20),它是BIA生物合成途径中的重要中间体。来自黄素腺嘌呤二核苷酸(FAD)依赖性氧化酶家族的小檗碱桥酶(BBE)催化7转化为四环小檗碱骨架(S)-可克劳林(8)(21)。最后,9OMT(22)、CYP719A(23, 24)和STOX(BBE样)(25)负责三个连续步骤——O-甲基化、亚甲基二氧桥形成和四电子氧化反应——将8转化为小檗碱(11)(图1A)。随着小檗碱生物合成的阐明,最近的研究重点转向了代谢工程和体外环境可持续生产。尽管已经实现了小檗碱的全新微生物合成,但由于催化效率和酶的多功能性限制,目前的产量尚未达到工业生产的要求(26, 27)。寻找替代生物合成酶的研究结果将为实现高效生产BIA提供重要的遗传元素。
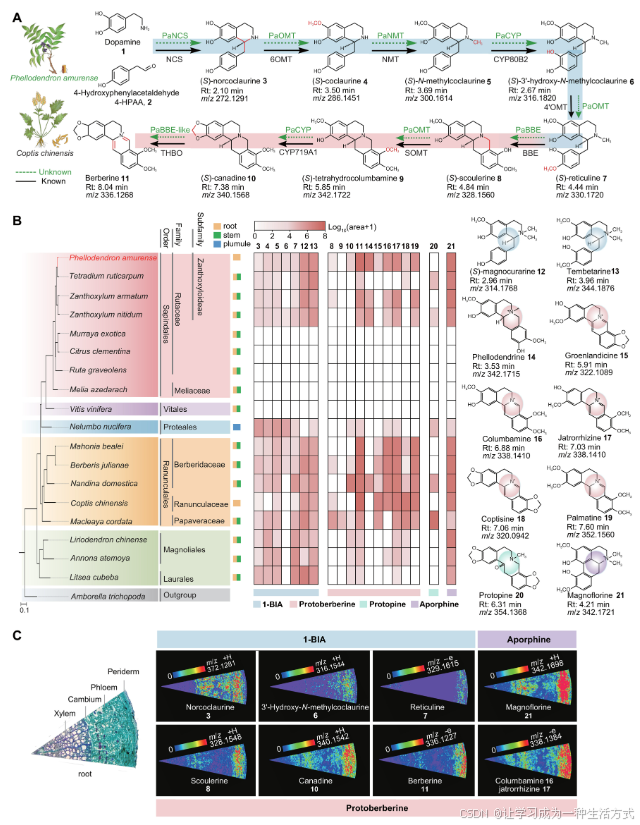
图 1. 黄柏属(P. amurense)中产生的原小檗碱和阿佛品类BIA。 (A) 小檗碱(11)的生物合成途径。黑色箭头表示来自小檗属的已报道酶,虚线箭头表示这些步骤在黄柏属中仍不明确。1,多巴胺;2,4HPAA;3,(S)-去氢可克劳林;4,(S)-可克劳林;5,(S)-N-甲基可克劳林;6,(S)-3′-羟基-N-甲基可克劳林;7,(S)-网状碱;8,(S)-可克劳林;9,(S)-四氢哥伦比亚碱;10,(S)-加拿丁;11,小檗碱。Rt,保留时间。 (B) 测试物种的系统发育树和BIA代谢物谱。该系统发育树是基于黄柏属(P. amurense)和其他18个物种的转录组或基因组数据(表S1)使用单拷贝直系同源基因构建的。代谢物是从17个物种的根部(黄色)和/或茎部(绿色)提取的,还包括莲子(N. nucifera)的莲胚(蓝色)。热图显示BIA水平,通过log10(峰面积+1)的值表示。BIA类型:1-BIA,原小檗碱、前小檗碱和阿佛品类。12,(S)-木兰啡;13,天贝他林;15,黄柏碱;15,格尔兰啡;16,哥伦比亚碱;17,贾托里赫辛;18,小檗啡;19,棕榈碱;20,前小檗碱;21,大花啡。 (C) 显微图像显示黄柏属(P. amurense)根部的横截面,MALDI-MSI图像生成的热图显示了代谢物的积累模式。
尽管在小檗属物种中已明确了解上述的生物合成途径,但是否在远缘关系的黄柏属物种中,小檗碱的丰富物种也遵循相同的途径仍不清楚。基于黄柏属和小檗属之间不同的谱系,我们假设黄柏属物种可能已经进化出不同的机制来合成小檗碱和其他BIA。为了验证这一假设,我们采用了基因组组装、转录组学、诱变、基因组比较、系统发育分析、蛋白质结晶、酶学测定和代谢组学的综合方法,解析了黄柏属(P. amurense)中小檗碱的生物合成途径。所得数据不仅确定了该谱系中小檗碱的六个生物合成步骤,还揭示了先前未知的酶学功能和机制。来自黄柏属的OMT酶通过不同的机制催化BIA中间体的N-甲基化。与小檗属中的FAD依赖性BBE不同,已鉴定出谱系特异性的CYP450酶催化黄柏属中的小檗碱桥形成。这些发现揭示了小檗碱的生物合成在小檗属和黄柏属中经历了不同的进化。小檗碱生物合成的分歧扩展了我们对植物BIA代谢的理解,并为代谢工程提供了重要的见解。
结果
黄柏属(P. amurense)中小檗碱的丰度
为了了解黄柏属(P. amurense)和其他17个植物物种在六个不同目(木兰目、月桂目、毛茛目、木兰目、葡萄目和无患子目)中的BIA分布,完成了液相色谱-质谱(LC-MS)分析,以概述各种组织中的19种BIA代谢物(表S1)。结果显示,这些BIA主要在木兰类植物(木兰目和月桂目)和早期分支的真双子叶植物中被检测到。数据还揭示了原小檗碱类和阿佛品类BIA,尤其是小檗碱(11,[M + H]+质量/电荷比(m/z)336.1268)、黄柏碱(14,[M + H]+ m/z 342.1715)、哥伦比亚碱(16,[M + H]+ m/z 338.1410)、贾托里赫辛(17,[M + H]+ m/z 338.1410)、棕榈碱(19,[M + H]+ m/z 352.1560)和大花啡(21,[M + H]+ m/z 342.1721),在黄木科物种(如黄柏、粗榄、刺木和光木)根部和/或茎部中高浓度积累(图1B及图S1和S2)。与这四个物种不同,其他无患子目物种和葡萄(Vitis vinifera)中未检测到BIA。在毛茛目中,像小檗属(如马赫尔·比尔利、黄柏朱利安、南丁丁和中国小檗等)也积累了高水平的原小檗碱类BIA,这与黄柏属相似(图1B)。这些发现证实了原小檗碱类BIA,特别是小檗碱,在远缘的无患子目(黄柏属)和毛茛目(小檗属)物种中具有独立和谱系特异性的分布。小檗碱的生物合成途径,尤其是在小檗属中,已经得到了完全阐明(图1A)。基于黄柏属和小檗属BIA代谢谱的相似性,我们提出这两种植物可能共享类似的小檗碱生物合成步骤。此外,完成了基质辅助激光解吸/电离质谱成像(MALDI-MSI),用于空间定位黄柏属(P. amurense)根部的BIA。所得图像显示,小檗碱途径代谢物和小檗碱衍生物定位于根部的外皮和韧皮部(图1C及图S3)。
基因组组装和P. amurense的全基因组重复倍增(WGD)
为了识别P. amurense中负责小檗碱生物合成的关键基因,我们生成了137.52 Gb的Illumina测序数据、73.14 Gb的PacBio Sequel II长读长测序数据,以及324.14 Gb的Hi-C配对末端读数,基于这些数据组装了染色体级别的单倍型基因组(图2A、图S4和表S2)。基因组调查、初步组装和染色体锚定分析构建了38条假染色体,覆盖了2.81 Gb的基因组大小,支架N50长度为76.63 Mb(图S5和表S3、S4)。该基因组的Benchmarking Universal Single-Copy Orthologs(BUSCO)完整性值达到了97.7%(表S5)。此外,P. amurense基因组中约87.03%由可转移元件组成,其中长末端重复反转录转座子(LTR-RTs)是最丰富的重复序列类别(表S6)。在P. amurense基因组中共预测了41,414个编码蛋白的基因,BUSCO完整性为97.60%,这表明基因组组装和基因预测的高完整性(表S5)。系统基因组学和时间推断分析表明,两种产生BIA的Zanthoxyloideae物种(P. amurense和Z. armatum)与未积累BIA的Aurantioideae和Rutoideae物种为姊妹关系(图2B和图S6),P. amurense与Z. armatum的分歧时间为2490万年前(Ma),95%置信区间(CI)为1833万至3153万年前(图S6)。
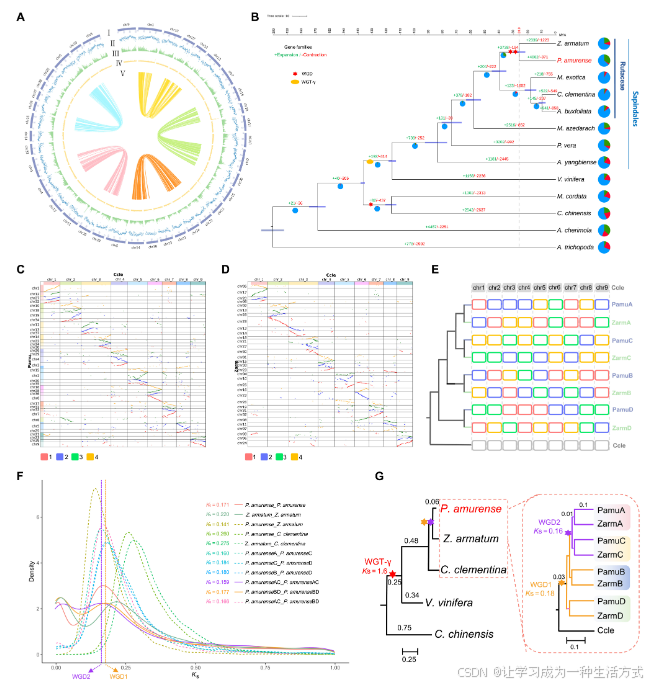
图2. P. amurense基因组的基因组特征、系统发育树和全基因组重复倍增(WGD)事件
(A) P. amurense的基因组特征。最外圈到最内圈的circos图展示了38条假染色体(I),基因密度(每Mb基因数)(II),1 Mb窗口中的GC含量(III),重复序列的密度(IV),以及circos图中心的每条连接线表示一对同源基因(V)。 (B) 基于P. amurense和12个候选物种的单拷贝直系同源基因构建的系统发育树。每个分支线上的绿色和红色数字分别表示扩展和收缩的基因家族数目。饼图展示了经历了基因家族收缩(红色)或扩展(绿色)的比例。分歧时间及其95%置信区间(CI)在分支节点上通过紫色水平条表示。WGT-γ表示gamma三重倍增事件。 (C) P. amurense和C. clementina基因组之间的共线性。以C. clementina基因组为参考,识别P. amurense基因组中每条染色体的同源基因对和共线性区块。 (D) Z. armatum和C. clementina染色体之间的共线性。 (E) 根据候选物种与C. clementina染色体(Chr1到Chr9)之间的共线性图谱,分别使用P. amurense子基因组和Z. armatum子基因组中的同源基因构建的系统发育树。P. amurense和Z. armatum中的红色、蓝色、绿色和橙色框代表与C. clementina的四种类型的共线性区块。 (F) P. amurense、Z. armatum、P. amurense子基因组、Z. armatum子基因组和C. clementina之间的直系同源基因和旁系同源基因的KS分布。 (G) P. amurense和Z. armatum的预测WGD事件。WGD1和WGD2分别表示P. amurense和Z. armatum基因组之间共享的WGD事件。
基因组内共线性分析揭示了P. amurense基因组中至少存在两次全基因组重复倍增(WGD)事件的遗迹(图2A和图S7、S8)。基因组间的共线性分析表明,P. amurense和Z. armatum基因组中的四个旁系同源段与Citrus clementina基因组中的一个直系同源区域相对应(图2C和D),这表明P. amurense和Z. armatum基因组中的两轮WGD事件可能发生在与C. clementina基因组分歧之后。P. amurense和Z. armatum的基因组分别映射到C. clementina基因组中。该映射在系统发育上将它们分为四个子基因组(PamuA、PamuB、PamuC和PamuD;以及ZarmA、ZarmB、ZarmC和ZarmD;图2E和图S9)。为了进一步探讨P. amurense WGD的系统发育位置,通过成对的子基因组对旁系同源基因的Ks(同义替代每个同义位点的替代数)分布分析显示了两个经过调整速率的Ks峰值:PamuAC旁系同源基因的0.16和PamuBD旁系同源基因的0.18。这一结果确认了P. amurense和Z. armatum在分裂之前共享了两轮WGD事件(图2F和G,及图S10)。这些结果表明,Rutaceae物种中可能发生了广泛的WGD事件和复杂的染色体进化。
小檗碱生物合成途径的阐明
为了识别参与P. amurense中小檗碱生物合成的基因,我们将先前对18个组织的转录组测序数据与小檗碱积累模式结合,进行了Pearson相关系数(r)分析。结果显示了与小檗碱产生相关的高排名候选基因(r > 0.80)。这些基因包括两个PaOMTs基因、一个PaNMT基因和四个PaCYPs基因,这些基因被选为功能分析的对象(图3A)。
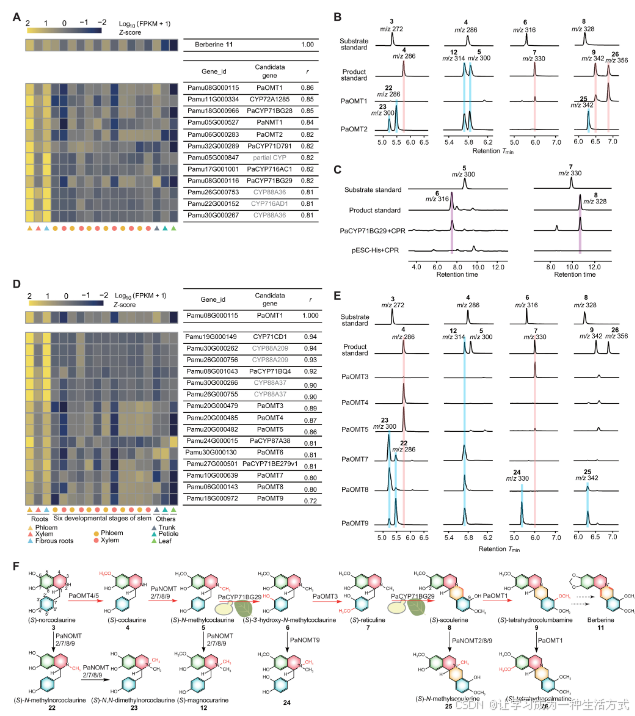
图3. 小檗碱生物合成途径的阐明
(A) P. amurense转录组数据与小檗碱含量之间的Pearson相关系数(r > 0.80)。热图展示了从log10(FPKM + 1)计算的z分数。灰色突出显示的CYP450s是未成功克隆的部分基因或报告的limonin生物合成同源基因(图S21)。FPKM表示每百万个比对片段中的每千碱基外显子模型的片段数。 (B) 使用3、4、6和8作为底物时,由纯化的PaOMT1和PaOMT2催化的化合物的提取离子色谱图。 (C) 使用5和7作为底物时,由PaCYP71BG29微体催化的化合物的提取离子色谱图。 (D) P. amurense转录组数据与PaOMT1基因之间的Pearson相关系数。灰色突出显示的基因是未成功克隆的或报告的limonin生物合成同源基因。 (E) 使用3、4、6和8作为底物时,由纯化的PaOMT4/5/7/8/9催化的化合物的提取离子色谱图。 (F) 本图总结了本文报告中验证的催化步骤。PaCYP71BG29的催化活性通过酵母和烟草表达系统进行了双重确认。N-甲基化的OMT成员被重新命名为NOMTs。Tmin为分钟时间。
首先,为了确定参与小檗碱生物合成的O-和N-甲基化反应,我们诱导重组的PaOMT1、PaOMT2和PaNMT1进行体外甲基化活性测试,使用了四种底物:(S)-norcoclaurine (3)、(S)-coclaurine (4)、(S)-3′-hydroxy-N-methylcoclaurine (6) 和 (S)-scoulerine (8)。这些测试确定PaOMT1和PaOMT2具有甲基化活性,而PaNMT1则没有。PaOMT1催化8号底物C-2和C-8羟基的连续O-甲基化,生成(S)-tetrahydrocolumbamine (9)(m/z 342)和(S)-tetrahydropalmatine (26)(m/z 356),其功能与C. chinensis中的Cc9OMT相同(图3B和图S11)。意外的是,PaOMT2没有表现出任何O-甲基化功能,但出乎意料地表现出对4号底物的N-甲基化活性,分别生成(S)-N-methylcoclaurine (5, m/z 300)和(S)-magnocurarine (12, m/z 314),通过单一和双重N-甲基化反应,这与C. chinensis中的CcNMT的催化活性相当(图3B和图S12)。此外,PaOMT2还催化了3、4和8号中间体上未饱和NH或NCH3基团的N-甲基化和N-去甲基化,表现出明显的底物宽容性(图3F)。这些此前未识别的活性增进了对BIA生物合成中N-甲基化的理解。
其次,使用CYP蛋白测试了BIA的氧化反应。根据已有报告,CYP450酶CYP80B1/2(24, 28, 29)、CYP80G1(29, 30)和CYP719A1(24, 31)分别催化5号底物的C-3′羟基化、7号底物的C-C苯基偶联反应以及(S)-tetrahydrocolumbamine (9)的美克伦二氧桥形成,四种候选PaCYPs—PaCYP71BG28、PaCYP71BG29、PaCYP71D791和PaCYP716AC1—在体外进行了催化功能研究,底物为5、7和9。结果表明,PaCYP71BG29对5和7的催化活性被检测到,而其他酶的活性没有被检测到(图S13)。PaCYP71BG29展示了在底物5的C-3′位上的羟化酶活性,生成6(m/z 316)(图3C和图S14)。此外,PaCYP71BG29还催化了7的黄连桥形成,生成8(m/z 328)(图3C和图S15),这与Coptis spp.中FAD依赖的氧化酶BBE的活性相同。此外,在Nicotiana benthamiana中瞬时表达PaCYP71BG29也确认了其对5的羟化活性(图S16)和对7的黄连桥形成活性(图S17)。
最后,通过使用PaOMT1作为诱饵基因进行共表达分析,阐明了小檗碱在黄檗属中的剩余生物合成步骤,选择了七个OMTs(PaOMT3到PaOMT9)和三个CYPs(PaCYP71BQ4、PaCYP71BE279v1和PaCYP87A38),这些基因的表达与PaOMT1高度相似(图3D)。体外生化实验显示了三种类型的甲基化活性。(i) 如PaOMT2测试所述,PaOMT7、PaOMT8和PaOMT9催化了N-甲基化活性,并具有广泛的底物宽容性(图3E和图S12)。(ii) PaOMT4和PaOMT5催化了3号底物C-6羟基的甲基化,生成4(图3E和图S18)。(iii) PaOMT3催化了6号底物C-4′羟基的甲基化,生成7(图3E和图S19)。此外,在不同底物的催化实验中,未检测到PaOMT6和三种PaCYP450s的活性(图S13和S20)。总之,这些生化数据阐明了黄檗属小檗碱途径中的6个酶催化步骤(图3F)。
PaNOMT9的晶体结构和催化机制
根据之前未识别的催化特性,N-甲基化OMT成员已重新命名为NOMT组(图3F)。为了理解PaOMT在P. amurense中催化的N-甲基化机制,我们解析了三元复合物PaNOMT9/S-腺苷甲硫氨酸(SAM)/4的晶体结构(蛋白质数据银行(PDB)ID:8ZN0),分辨率为1.8 Å(图4A)。PaNOMT9的蛋白质结构形成了二聚体(标记为A和B)。这两个不同的单体分别构成N端二聚体化域(1到165氨基酸,主要由α1到α10、β1到β3和η1组成)和C端Rossmann折叠的SAM/S-腺苷高半胱氨酸(SAH)结合域(199到359氨基酸,α12到α16、β4到β10和η2),两者通过α11螺旋(166到198氨基酸)连接(图4A)。在PaNOMT9的二聚体化域(α6和α10)、SAM/SAH结合域(α15和η2)以及α11螺旋的界面处,17个残基距离底物4在5 Å范围内(图S22A和S23)。蛋白质-配体相互作用分析揭示,H127与C6-甲氧基和C7-羟基之间形成氢键,而Y159与NH基团之间形成氢键(图4B)。核心底物4被F123、F126、W262、I265、M312、I315、G316、V320和L325等疏水性残基环绕,并与这些残基相互作用。此外,距离活性位点较远的D185、D293和L295通过序列比对被鉴定为N-甲基化和O-甲基化OMT蛋白的差异残基(图4C和图S22B及S23)。
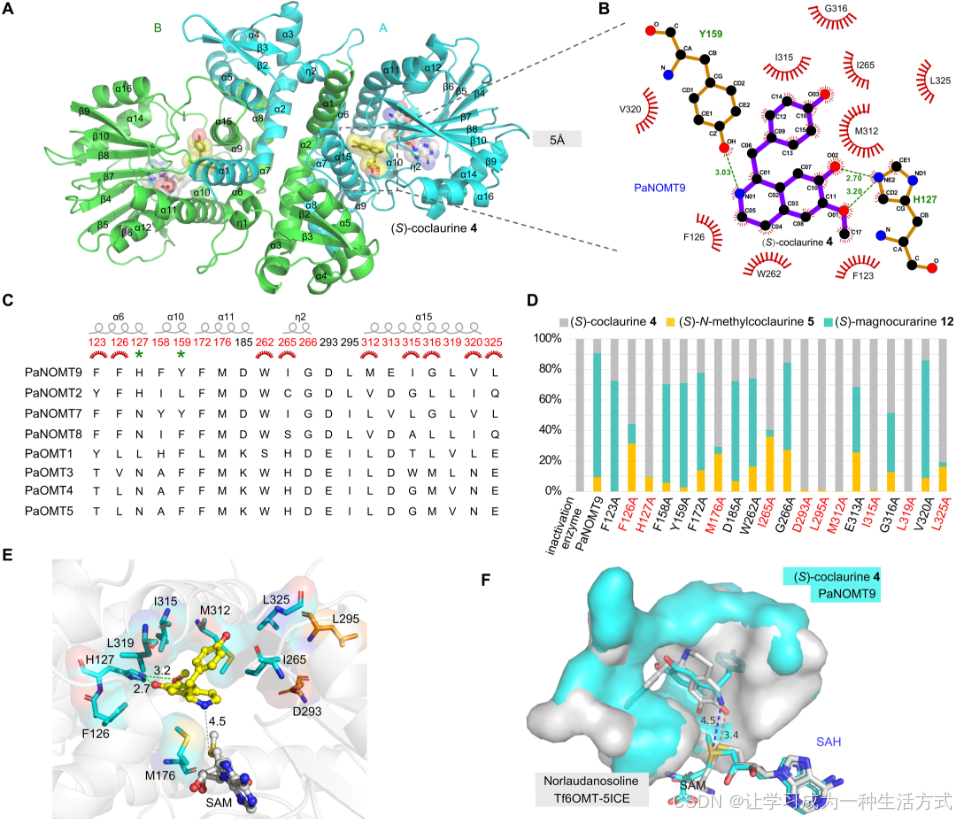
图4. PaNOMT9的晶体结构和催化机制
(A) PaNOMT9/4/SAM三元复合物的晶体结构总体视图。 (B) PaNOMT9与配体4之间的蛋白质-配体相互作用图。红色标记表示疏水性残基。 (C) N-甲基化活性OMT蛋白PaNOMT9/2/7/8与O-甲基化OMT蛋白PaOMT1/3/4/5之间的氨基酸序列比对。 (D) PaNOMT9及其20个通过定向突变得到的变体的催化活性,以堆叠条形图表示。 (E) 由PaNOMT9/4/SAM形成的三元复合物的晶体结构,其中突变这些关键残基会降低催化活性。用青色突出显示的残基表示这些残基与底物之间的距离小于5 Å,而用橙色表示的残基超出了这一范围。灰色虚线表示4的N原子与SAM甲基化基团之间的距离为4.5 Å。 (F) 两个三元复合物晶体结构的叠加:Tf6OMT/norlaudanosoline/SAH和PaNOMT9/4/SAM。这两个三元复合物的蛋白质-配体结合构象差异导致了O-/N-甲基化功能的差异。
上述PaNOMT9的20个残基被逐一用丙氨酸(A)替代,生成了20个变体,然后测试了每个变体以4为底物的活性。反应中甲基化产物(5)或二甲基化产物(12)的含量通过LC-MS分析估算(图4D)。一方面,结果表明,M312、I315和L319的丙氨酸替换几乎完全废除了N-甲基化活性(图4D)。另一方面,四个变体—F126A、M176A、I265A和L325A—表现出明显高于野生型的5/12(甲基化/二甲基化)比值(图4D)。这些结果表明,疏水相互作用对于底物结合口袋的构建至关重要(图4E)。此外,位于远离活性位点的D293A和L295突变体几乎完全失去了N-甲基化活性,这可能归因于蛋白质结构的扰动。PaNOMT9的H127A变体只能催化无效的N-甲基化反应,表明H127与底物4之间的氢键对于底物结合构象的形成至关重要(图4D和E)。我们进一步生成了11个额外的H127突变体,结果表明,保守替换(H127N)和具有相似性质的替换(H127K和H127R)保留了对4的野生型N-甲基化活性。相比之下,H127P突变体几乎完全失去了N-甲基化活性(图S24A)。H127A和H127P突变体,位于远离C6-甲氧基和C7-羟基的地方,导致活性口袋内出现较大的空腔,这表明H127在底物结合构象中起着关键作用(图S24B)。
为了理解PaNOMT9催化的N-甲基化机制,我们对晶体结构进行了比较。已报道的来自Thalictrum flavum的Tf6OMT(PDB ID:5ICE)和来自Coptis japonica的CjCNMT(PDB ID:6GKV)分别催化O-甲基化和N-甲基化,并已详细解析了它们的晶体结构(32, 33)。基于这些数据,我们将Tf6OMT和CjCNMT的蛋白质结构与PaNOMT9进行了比较(图S22)。一方面,Tf6OMT和PaNOMT9的整体结构相似,它们的SAM和SAH位置相对接近;然而,这两种结构有明显不同的蛋白质折叠(图S22C)。将Tf6OMT/norlaudanosoline/SAH和PaNOMT9/4/SAM的复合物结构叠加,以比较它们的活性口袋和底物结合构象(图4F和图S22C)。构象比较揭示,Tf6OMT和PaNOMT9的活性位点口袋中的底物定向不同(图4F)。底物4的NH基团与SAM供体的距离相对较近(4.5 Å),而在Tf6OMT复合物中,C-6 OH基团则与SAM供体的距离较近(3.4 Å)。此外,与PaNOMT9和底物3、6、8的对接模拟展示了与4相似的构象(图S25)。另一方面,与CjCNMT(PDB ID:6GKV)的蛋白质结构相比,非同源的PaNOMT9呈现出完全不同的整体结构,包括SAM和底物结合区域、氢键和疏水性残基的构象,尽管这两种蛋白质展现出相似的N-甲基化活性。这些结果表明,PaNOMT9在这些区域及其周围的构象灵活性和可塑性使得底物能够进行N-甲基化。
PaCYP71BG29在小檗碱桥形成中的新功能化和催化机制
通过同源基因挖掘、系统发育分析和功能实验,追溯了PaCYP71BG29催化的小檗碱桥形成的起源和进化。系统发育分析表明,来自P. amurense的PaCYP71BG29和另外五个基因、Z. armatum的九个基因以及C. clementina的两个基因与水稻的OsCYP71P1(序列相似度为46%到58%)聚类在同一分支(图5A、图S26和表S7、S8)。进一步与氨基酸序列的比较显示,PaCYP71BG29与OsCYP71P1(一个色胺5-羟化酶(T5H))具有47%的相似度(34)。
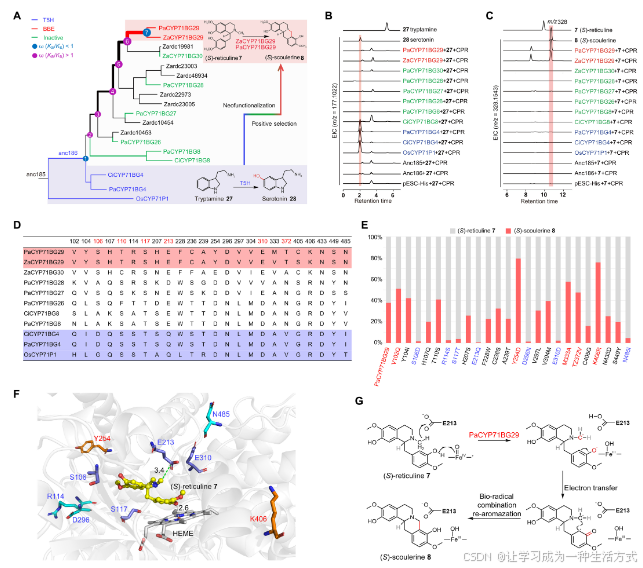
图5. PaCYP71BG29的新功能化及其催化机制的确定
(A) 基于P. amurense、Z. armatum和C. clementina中PaCYP71BG亚家族成员构建的最大似然系统发育树。OsCYP71P1作为外群。Ka/Ks是非同义(Ka)与同义(Ks)替代率。 (B) 和(C) 提取离子色谱图(EIC),表征由候选CYP微粒体和两个底物——色胺27和(S)-网状碱7组成的酶促反应生成的化合物。 (D) PaCYP71BG29、ZaCYP71BG29和其他CYP71BG亚家族成员的氨基酸序列比对。展示了24个氨基酸,其中六个位点(红色标记)与7相距5 Å以内。 (E) 堆叠条形图展示了PaCYP71BG29及其24个变体对两个底物的催化偏好:(S)-网状碱7和(S)-小檗碱8。 (F) PaCYP71BG29/HEME/7的分子对接模型,显示了那些关键残基,它们增强(红色)或降低(蓝色)催化活性。底物7与关键残基E213和HEME之间的距离分别为3.4和2.6 Å。 (G) 提出了由PaCYP71BG29催化的小檗碱桥形成的机制。
CYP71BG亚家族成员的系统发育树被分为七个节点(节点1至节点7)。根据系统发育树,选择了九个CYP71BG基因及其两个祖先基因(anc185和anc186)来诱导重组蛋白进行活性测试(图5B和C)。首先,根据OsCYP71P1的催化活性,使用色胺(27)作为底物进行体外酶促实验,并选择OsCYP71P1作为阳性对照。结果显示,PaCYP71BG4、CiCYP71BG4和OsCYP71P1将色胺27转化为血清素28(图5B和图S27)。此外,实验表明,祖先CYP71BG基因anc186也编码T5H,尽管其催化活性较为温和。相反,P. amurense和Z. armatum基因组中从基因扩展产生的节点2中的CYP719BG候选基因对色胺27没有羟基化活性(图5A和B)。 其次,选择网状碱7作为底物测试小檗碱桥活性。催化实验显示,节点7中的PaCYP71BG29和ZaCYP71BG29表现出了小檗碱桥活性,将(S)-网状碱7转化为(S)-小檗碱8(图5A和C)。相比之下,P. amurense和Z. armatum中其他重复的CYP71BG基因无法催化7到8的转化。这些结果表明,PaCYP71BG29和ZaCYP71BG29在植物进化过程中发生了新功能化和分化。 最后,使用双比率分支模型检测了正向选择。结果显示,从节点2到节点6的分支ω值大于1。这些值还表明,PaCYP71BG29和ZaCYP71BG29发生了新功能化。获得的催化作用负责P. amurense和Z. armatum中小檗碱桥的形成(图5A)。
为了理解PaCYP71BG29在小檗碱生物合成中的环化酶活性机制,使用(S)-网状碱7作为配体,通过AlphaFold2完成了对接模拟(图S28)。通过序列比对进一步选择了在活跃与不活跃CYP71BG酶中显著不同的残基(图5D)。通过比对选择了24个不同的残基,其中六个残基,包括S106、T110、S117、E203、E310和T372,定位在7周围5 Å以内(图5D和图S29)。这些PaCYP71BG29的残基接受了一系列的突变,以研究其对T5H功能的贡献,并测试了使用7作为底物的突变体的活性(进行三次重复实验;图5E)。几个突变体增强了小檗碱桥的形成活性;例如,PaCYP71BG29的突变体Y254D和K406R表现出明显有效的转化率,分别为79.4%和75.9%,高于野生型的37.8%。PaCYP71BG29的七个突变体,包括S106D、R114S、S117T、E213Q、D296N、E310D和N485I,几乎失去了小檗碱桥形成活性,表明它们在将7转化为8中的关键作用(图5E)。这些变体中的S106D、S117T、E213Q和E310D位于5 Å范围内,可能会影响底物结合。另一方面,R114S、D296N和N485I这些位于远离活性中心的变体,可能通过改变蛋白质构象和表面结构来影响PaCYP71BG29的催化活性(图5F)。基于这些数据,我们提出这些残基的变化导致了CYP71BG29的新功能化。
PaCYP71BG29/HEME/7的对接模型显示,关键残基E213的羧基与7的N-甲基基团接近(3.4 Å),而HEME铁中心则靠近7的C3′-OH基团(2.6 Å)(图5F)。进一步参考已报告的BBE(35),提出了PaCYP71BG29在小檗碱桥形成中的催化机制:HEME辅因子去质子化C3′-OH基团,增强其亲核性,来自N-甲基基团的氢化物转移到E213,反应中间体的再芳构化导致8的环化反应(图5G)。因此,尽管迄今为止所有检查过的Ranunculales植物都使用FAD依赖性氧化酶催化小檗碱桥反应,但Rutaceae科的成员(P. amurense和Z. armatum)可能通过趋同进化出一个独特的CYP450成员,通过类似的化学机制来介导相同的氧化反应。
讨论
尽管小檗碱的生物合成已经在小檗属植物中得到阐明,但是否其他谱系植物也存在相同的生物合成途径仍然不清楚。对此问题的回答不仅有助于理解小檗碱的生物合成,还能帮助我们了解不同植物中的多样化代谢途径。已报道的小檗属植物中小檗碱的生物合成包括一个NCS、三个OMTs(6OMT、4′OMT和9OMT)、一个NMT(CNMT)、两个CYPs(CYP80B和CYP719A)、一个BBE和一个类似BBE的酶(THBO)。在P. amurense基因组中对同源基因的注释显示,与小檗属植物中的基因相比,NCS(33%到37%)、OMTs(26%到49%)、NMT(33%到37%)、CYPs(23%到42%)和BBE基因(29%到57%)的身份非常低(表S9)。此外,在P. amurense中未发现CYP80和CYP719基因。因此,这些特征表明,小檗碱的生物合成途径在P. amurense和小檗属植物之间可能存在差异。此外,基于这些特征,我们假设在P. amurense的小檗碱生物合成中可能涉及未知的甲基化和氧化反应机制。以下将通过OMT的N-甲基化和CYP酶的新功能化来验证这一假设。
本文报告的PaNOMT9的催化活性揭示了一个涉及小檗碱生物合成的双重甲基化机制。SAM依赖的MTs在植物天然产物多样化中起着重要作用。以往的研究已表征了将甲基转移到化合物的S、N、O或C原子上的MT成员,生成甲基化的天然产物(36)。根据S、N、O或C的修饰,MTs已被分类为SMT、NMT、OMT和CMT。迄今为止,BIA生物合成中的MT特异性表明,OMT和NMT成员严格地将甲基转移到C-OH和NH基团上(37)。与此相反,已有报告表明OMT家族的某些II类和III类成员在黄嘌呤和邻苯二甲酸中都具有O和N-甲基化活性(表S10)(38, 39)。在此,我们的研究数据增加了MT的另一个亚类,NOMTs。来自黄柏(Phellodendron)的PaNOMT2/7/8/9与Coptis的CNMT基因具有相似的催化活性,是首个能够催化BIA生物合成中N-甲基化的OMT成员。来自P. amurense的与小檗碱生物合成相关的NOMT基因(PaNOMT2/7/8/9)与OMT家族II类亚家族呈姊妹群(图S30A)。NOMT基因的系统发育分支包含来自P. amurense和Z. armatum的大量串联重复基因,且仅在柑橘中观察到一个同源基因(CiOMT2),与PaNOMT2/7/8/9的序列同源性为64%到66%(图S30B和S31)。我们的催化实验表明,CiOMT2不能对底物4执行N-甲基化活性(图S30B),表明N-甲基化功能的获得是在它们与柑橘的分离后演化而来的。PaNOMT9的N-甲基化催化机制与已报道的BIA OMT或NMT酶有所不同,提供了有价值的信息,有助于加深对BIA结构甲基化的理解,并推动潜在BIA药物的代谢工程。
本文报告的PaCYP71BG29的小檗碱桥形成活性揭示了植物专用代谢产物生物合成中来自非同源酶的相同催化活性,这是一个先前未曾发现的案例。在以往的研究中,Ranunculales超家族中的FAD依赖性氧化酶BBE,如小檗、罂粟、千屈菜和小檗属等植物的BBE在小檗碱生物合成中的小檗碱桥形成中起着保守作用(21, 40-42)。然而,来自P. amurense基因组的PaBBE基因仅有一个,与Ranunculales植物的BBE聚成同一分支。此外,基因表达分析显示,PaBBE在所有测试的P. amurense组织中都是沉默的(图S32)。这一数据表明,这种药用植物中可能存在未知的机制。为了验证这一点,我们挖掘了基因组和转录组序列,揭示了与小檗碱积累模式相关的CYP71BG成员。比较基因组分析发现,一方面,Ranunculales基因组中没有注释CYP71BG成员;另一方面,这些基因来自基因组扩展。这些数据为PaCYP71BG29催化的小檗碱桥形成和生物合成步骤提供了证据,表明这可能是P. amurense独立进化的结果。此外,我们的生化实验显示,CYP71BG酶参与了与植物发育和应激反应相关的植物胺血清素和褪黑激素的生物合成。系统发育分析表明,CYP71BG成员的明显扩展和正向选择导致了Phellodendron中CYP71BG29的新功能化。
9OMT(PaOMT1)和BBE(CYP71BG29)在黄柏染色体8上形成一个生物合成基因簇,该基因簇进一步经历了WGD1和WGD2事件,扩展了N-甲基化的PaNOMT7和PaNOMT9(图S33),这与小檗属植物中小檗碱生物合成基因的分散分布不同。此外,4′OMT(PaOMT3)和6OMT(PaOMT4和PaOMT5)在染色体20上聚集在一起,WGD事件也加速了共线性OMT的扩展。我们还发现,小檗碱生物合成基因在黄柏和花椒中是保守的,CYP450和OMT基因的重复和新功能化发生在它们与柑橘分开之后(图S33)。小檗碱生物合成基因簇及其重复事件扩展了我们对代谢基因簇形成和独立进化的理解(43)。
植物专用代谢物被视为复杂性状,可以通过研究其进化轨迹来理解(44)。相同的代谢特征经常在远缘物种中独立进化出现,包括平行进化和趋同进化。例如,金丝菊和大麻中由同源酶催化的大麻素生物合成的平行进化(45)。尽管多项研究已报道了咖啡因(39)、藏红花色素(46)和稻米乳酸酮(47)生物合成的趋同进化,但它们在不同谱系中的生物合成酶可能具有同源关系。参照平行进化和趋同进化的区别(48),我们的研究展示了一个经典的趋同进化案例,在两个远缘谱系中——小檗属和黄柏,两个完全非同源的酶和不同的蛋白质折叠,尤其是具有NMT催化功能的OMT基因和具有小檗碱桥酶活性的CYP450基因。这些数据表明,酶的新功能化是推动结构修饰和多样化的驱动力。
总之,我们报告了黄柏的高质量基因组,并解密了一个替代的小檗碱生物合成途径,特别是识别了N-甲基化OMT基因和由CYP450催化的小檗碱桥活性。所识别的小檗碱生物合成步骤来源于WGD事件和新功能化。这些发现揭示了小檗属和黄柏物种中小檗碱生物合成的趋同进化模型。此外,我们的研究增强了我们对小檗碱生物合成酶的理解,为工程化具有多种药理活性的复杂BIA药物提供了支持。
材料与方法
植物材料与化学标准
黄柏(P. amurense)植物生长在中国哈尔滨东北林业大学的研究站内。新鲜的黄柏叶片被收集后迅速放入液氮中,并储存于-80°C冰箱中以供基因组测序。新鲜的黄柏根部被收集并分成两个部分,一个部分在液氮中冷冻用于代谢物提取,另一个部分用于MSI(MALDI-MSI)分析。其他16种植物的根和/或茎,以及荷花(N. nucifera)的干莲子芽也被收集用于代谢分析(表S1)。所有化学标准如表S11所列,均从不同公司购买。
代谢物的LC-MS分析
所有样品(含三次重复)在50°C下烘干后,研磨成细粉,并通过60目筛过滤。称取200 mg细粉放入15 ml玻璃管中,加入10 ml甲醇:水混合液(6:4,v/v),并漩涡混合1分钟。然后在室温下静置过夜,并在超声波清洗机(240 W、40 Hz)中超声处理40分钟。玻璃管在9000g下离心10分钟,收集上清液中的代谢物,转移到清洁管中用于代谢物分析。接着,超声清洗后的上清液再次在12,000g下离心10分钟,所得的干净上清液转移至1.5 ml管中,并通过0.22 μm滤膜过滤。
样品通过SHIMADZU UPLC-QTOF-MS系统进行检测(LCMS-9030),配有SHIMADZU GIST C18色谱柱(2 μm,2.1 mm x 100 mm)。流动相由0.1%甲酸水溶液(A)和乙腈(B)组成,线性梯度洗脱程序如下:0到1分钟,15% B;1到8分钟,15%到45% B;8到10分钟,45%到80% B;10到11分钟,80%到15% B;11到14分钟,15% B。流速为0.2 ml/min,柱温为40°C。质谱仪在数据依赖采集模式下操作,电喷雾电离(ESI)源在正离子模式下运行(m/z 100到700)。气体温度维持在400°C,气体流量为10升/分钟,碎片电压设置为120 V,碰撞能量为20到40 eV。
MALDI成像
黄柏的冷冻根部被包裹在5%的羧甲基纤维素(w/v)中,然后使用干冰将其固化为块状。冷冻根部在-20°C下被切割成30 μm厚的切片,切片立即粘附于涂有铟锡氧化物的玻片上以供后续成像分析。与此同时,将60 mg 2,5-二羟基苯甲酸(DHB)溶解在2 ml甲醇和水的混合液中(含0.1%三氟乙酸),比例为7:3(v/v)(49)。所得的DHB溶液用于正模式MALDI实验。所有测量使用与Orbitrap质谱仪(“Q Exactive”,Thermo Fisher Scientific,德国不来梅)耦合的AP-SMALDI离子源(“AP SMALDI10”,TransMIT GmbH,德国吉森)进行。简而言之,使用固态激光(λ = 343 nm),重复频率为2000 Hz,用于在正离子模式下分析样品。使用DHB矩阵增加电离效率。全扫描模式下的测量速率(扫描范围m/z 150到1400)约为1.5秒/像素,质量分辨率为70,000,分辨率对应m/z 200时的质量精度为2 ppm。MSI分析的空间分辨率为30 μm。数据分析使用MirionV3软件进行。
基因组测序、调查与组装
使用DNeasy Plant Mini Kit(QIAGEN,德国)从冷冻叶片中提取高质量基因组DNA样品。通过NanoDrop分光光度计(Thermo Fisher Scientific,美国)评估和测量DNA样品的质量和数量。DNA样品被消化为20-kb大小。然后,使用SMRTbell模板准备试剂盒构建DNA文库。简要而言,根据制造商的协议,DNA文库构建步骤包括DNA浓缩、损伤修复、末端修复、发夹接头连接和模板纯化。基因组测序的环状共识测序数据是在PacBio Sequel II平台(Pacific Biosciences,美国)上产生的,遵循制造商的协议。所获得的DNA序列用于下面描述的基因组组装。所有比较包括三次独立的生物学重复实验。 从冷冻叶片、茎和根中提取总RNA样品,使用RNAprep Pure Plant Kit(TIANGEN,中国)。随后,使用Qubit RNA Assay Kit和Agilent 2100 Bioanalyzer(Agilent Technologies)评估RNA样品的质量。RNA测序在Novogene使用Illumina HiSeq 2500平台进行,以生成150-bp配对末端读段。使用ProximoTM Hi-C Plant试剂盒从P. amurense的新鲜叶片中构建了Hi-C文库。cDNA文库在Illumina HiSeq平台上进行测序,生成150-bp的长读段。使用Illumina数据根据K-mer分布分析(K = 21)估算P. amurense的基因组大小。过滤PacBio Sequel读段以去除短的和低质量的读段。结果高质量的读段用于使用Hifisam(v0.19.5)进行de novo组装,采用默认参数(50)。使用Purge_dups(v1.2.5)(51)去除组装的contig序列的杂交。使用ALLHiC(v0.9.8)(52)根据Hi-C数据生成一个支持等位基因的染色体级基因组。
基因注释与质量评估
基因注释通过de novo预测、同源蛋白证据和EST证据方法完成。组装后,RNA测序(RNA-seq)数据通过HISAT2(v2.2.1)(53)映射到基因组上。对于ab initio预测,AUGUSTUS模型通过BRAKER2(v2.1.5)(54)使用RNA-seq数据训练。此外,结合同源证据的ab initio预测与MAKER管道结合,生成最终的基因模型。使用BUSCO(v5.2.2)(55)和embryophyta_odb10数据库评估基因组的完整性。注释后,使用Tandem Repeat Finder(v4.07)(56)预测串联重复。LTR_retriever(v2.6)(57)、LTR_FINDER(v1.0.6)(57)和RepeatModeler(v2.0)用于构建重复序列数据库。对于de novo预测,生成的LTR和通过RepeatModeler生成的共识重复文库被合并,并作为RepeatMasker的输入数据。
比较基因组分析
使用P. amurense和其他14个已测序植物物种(包括Z. armatum(58, 59)、Murraya exotica(60)、C. clementina(61)、Atalantia buxfoliata(62)、Melia azedarach(63)、Pistacia vera(64)、Acer yangbiense(65)、V. vinifera(66)、Macleaya cordata(67)、C. chinensis(15)、Annona cherimola(68)和一个外群物种[Amborella trichopoda;(69)]的同源基因组群进行系统发育分析。使用OrthoFinder(v2.3.11)(70)获得15个物种中的单拷贝家族,采用默认参数。通过RAxML(v8.0.17)(70)使用最大似然方法并进行1000次自举重复,构建了包含221个单拷贝同源基因的系统发育树。使用PAML(v4.9)(71)中的MCMCTREE估计15个物种之间的分歧时间,校准点来自TimeTree数据库(www.timetree.org)。校准点包括P. amurense和C. clementina之间的分歧(25到69百万年)、M. azedarach与P. vera(63到105百万年)、A. yangbiense与V. vinifera(110到124百万年)、A. cherimola与A. trichopoda(180到205百万年)。 使用CAFE(v4.2.1)(72)分析了基因家族的扩展与收缩。基于Gene Ontology和京都基因与基因组百科全书数据库,使用R包ClusterProfiler(v4.0)(73)分析了扩展基因家族的功能富集。
WGD分析
使用MCScan(Python版本)和默认参数建立基因组内和基因组间的同源区块。基于检测到的同源基因对识别同源区块,并使用Whole-Genome Duplication Identifier(WGDI)管道(v0.6.5)(74)估算共线基因之间的Ks值。此外,为了检测P. amurense基因组中可能的多倍体化遗迹,使用MCScan识别了P. amurense、Z. armatum和C. clementina基因组内的同源基因区块。基于C. clementina基因组提取每个区块的共线基因,并使用IQ-TREE2(v2.2.2.7)(75)构建共线基因树。基因树作为输入上传到ASTRAL(v5.6.1)(76),推断子基因组系统发育。最后,使用ksrates包(GitHub - VIB-PSB/ksrates: ksrates is a tool to position whole-genome duplications relative to speciation events using substitution-rate-adjusted mixed paralog-ortholog Ks distributions.)根据调整后的混合图追踪平行基因和同源基因的Ks分布,追踪WGD事件。
候选基因的鉴定与相关性分析
报告中的生物碱(BIA)生物合成基因,包括NCS、MTs和CYP450,被选作查询序列,并通过BLASTp在P. amurense、Z. armatum和C. clementina基因组中进行同源基因的鉴定,E值为1 × 10−10。简而言之,选用Papaver somniferum中的PsNCS1(AAX56303.1)和PsNCS2(AAX56304.1)、C. japonica中的CjNCS(BAF45338.2)、T. flavum中的TfNCS(ACO90248.1)作为查询序列,鉴定候选的NCS编码基因。P. somniferum中的Ps6OMT(AAQ01669.1)以及C. japonica中的Cj6OMT(BAB08004.1)、Cj4′OMT(BAB08005.1)和Cj9OMT(BAA06192.1)作为查询序列,用于鉴定OMTs的同源基因。C. japonica中的CjNMT(AB061863.1)作为查询序列,用于鉴定NMTs。C. japonica中的CjCYP719A1(BAB68769.1)、CjCYP80B2(BAB12433.1)、CjCYP80G2(BAF80448.1)、P. somniferum中的PsCYP80B1(AAF61400.1)和PsCYP719B1(ABR14720.1)、Berberis stolonifera中的BsCYP80A1(AAC48987.1)作为查询序列,根据命名规则鉴定候选的CYP450亚家族(身份比> 55%)。
基于P. amurense的转录组(PRJNA859281)和代谢组数据,利用候选基因的标准化表达水平与berberine的含量水平,分析了Pearson相关系数(r)。筛选出与候选MTs和CYP450s基因表现出高相关性(r > 0.8)的基因用于功能验证。然后,进一步选择功能基因作为诱饵,通过共表达分析选择额外的候选基因。
MTs基因在大肠杆菌中的异源表达与功能验证
候选的MT基因被克隆或合成,并克隆到pET-28a载体中(见表S12)。随后,将得到的阳性重组质粒导入大肠杆菌BL21(DE3)菌株。每个载体的阳性克隆都被选中并诱导重组蛋白的表达。开发了一种通用协议来表达每个候选基因并生产其重组蛋白。选中阳性克隆,接种20 ml含有50 μg/ml卡那霉素的液体Luria-Bertani(LB)培养基中,放置于250 rpm/min、37°C的摇床上。当细胞悬液的光密度(OD600)达到0.6时,加入异丙基-β-d-硫代半乳糖苷(IPTG;0.3 mM)以诱导蛋白表达。加入IPTG后,将培养箱温度设为16°C。培养过夜后,通过离心(8000g,5分钟,4°C)收获细胞。将沉淀重悬于1 ml含有50 mM tris-HCl缓冲液(pH 7.4)、10%(v/v)甘油和5 mMβ-巯基乙醇的溶液中。通过超声破碎冰上培养5分钟。将裂解液以8000g离心10分钟,收集上清液并存储于−20°C进行酶活性检测。
为了纯化候选的重组蛋白,增大了每个重组载体的大肠杆菌培养体积至100 ml LB培养基。其他条件保持一致。将沉淀重悬于30 ml裂解缓冲液(pH 8.0;50 mM NaH2PO4,30 mM NaCl和10 mM咪唑)中。提取步骤与小规模提取相同。得到的30 ml上清液加载到Ni-NDA柱(Ni-NDA Beads,Smart-Lifesciences,中国)上进行重组蛋白纯化。为结晶纯化进一步使用凝胶过滤色谱法(与150 mM NaCl和20 mM Hepes,pH 8.0平衡)进一步纯化蛋白,最终通过Amicon Ultra-15 Ultracel-30 K离心过滤器(Merck Millipore)浓缩蛋白。使用蛋白定量试剂盒(TransGen Biotech,中国)测定蛋白浓度,并通过SDS-聚丙烯酰胺凝胶电泳(SDS-PAGE)评估纯度。
MTs的酶活性在37°C、200 rpm摇动下的100 μl反应体系中进行过夜反应,反应体系包括Gly缓冲液(pH 9.0),10 μg纯化蛋白,2 mM SAM,底物(S)-norcoclaurine、(S)-coclaurine或(S)-scoulerine,浓度为50 μg/ml,或3′-hydroxy-N-methylcoclaurine,浓度为20 μg/ml,反应结束后加入50%甲醇(v/v)终止反应。选择Cc6OMT、Cc4′OMT、Cc9OMT、CcNMT作为阳性对照,空载体作为阴性对照。
通过超高效液相色谱-串联质谱(UPLC-MS/MS)检测反应样本。UPLC-MS/MS系统(SCIEX TripleTOF 6600+)配备三重四极体-线性离子阱质谱仪。简而言之,1 μl提取液注入后,使用Kinetex C18 100A分析柱(4.6 mm×150 mm,2.6 μm)进行分离,柱温设为35°C。流动相包括A相(0.1%甲酸)和B相(乙腈)。梯度程序如下:0至1分钟,10%B;1至10分钟,10至95%B;12.3至13分钟,95%B;13至15分钟,95至10%B,流速为0.5 ml/min。质谱仪工作在多重反应监测模式下,扫描时间为每个转移35 ms。质谱仪的参数设置如下:正离子模式ESI模式;喷雾电压为3.5 kV;喷雾温度为550°C;气帘气体为35 psi;GAS1为40 psi,GAS2为60 psi。
S. cerevisiae中CYP450基因的异源表达与功能验证
为了验证候选CYP450的活性,选择了转基因的酿酒酵母Saccharomyces cerevisiae WAT11(包含拟南芥细胞色素P450还原酶AtCPR1)作为异源表达宿主。候选CYP450基因被克隆或合成,并克隆到pESC-His载体中(引物见表S12),然后转入S. cerevisiae WAT11中。筛选阳性酵母菌落表达CYP450蛋白。经过培养和诱导后,酵母细胞被收获、破碎并进行差速离心,得到微粒体,具体操作参考之前的研究。
对于酶活性检测,选择表达pESC-His空载体的酵母作为阴性对照。反应体系包括200 μl微粒体、0.5 mM还原型烟酰胺腺嘌呤二核苷酸磷酸和1 μl底物(1 mg/ml),在30°C下孵育2小时。反应样本经过离心和过滤后,使用Agilent UPLC-QTOF-MS检测。UPLC-QTOF-MS系统配备ACQUITY UPLC HSS-T3 C18柱(1.8 μm,2.1 mm×100 mm),流动相包括水中0.1%甲酸(A)和乙腈(B),线性梯度洗脱程序如下:0至2分钟,5%B;2至10分钟,5至18%B;10至15分钟,18至95%B;15至17分钟,95%B。流速设为0.3 ml/min,柱温设为40°C。质谱仪使用Agilent 6430 QTOF,工作在正离子模式下,扫描范围为m/z 100到800,气体温度保持在350°C,气流速率为8升/分钟,碎片电压设置为120 V,碰撞能量为20 eV。
PaCYP71BG29在本氏烟草中的瞬时表达
为了进一步验证PaCYP71BG29在体内的功能活性,该基因被克隆至pEAQ-eGFP载体中,并转化至根癌农杆菌(Agrobacterium tumefaciens,GV3101)。阳性克隆在10 ml含有卡那霉素和利福平的LB培养基中培养。培养物以5000g离心5分钟,沉淀重悬于渗透缓冲液(10 mM MES缓冲液、10 mM MgCl₂和150 μM乙酰香豆素),并在28°C黑暗条件下孵育1小时。将Agrobacterium悬液(每个菌株OD600为0.3至0.6)渗透入5至6周龄的本氏烟草叶片。3天后,将底物5和7(50 μM,溶解于0.1%二甲基亚砜水溶液中)渗透入先前经根癌农杆菌渗透的叶片背面。1天后,收集叶片进行LC-MS/MS分析。生物学重复包括从三条不同基因型中收集的三至四片叶子。使用空载体渗透的叶片和无底物的候选基因作为对照。
PaNOMT9的结晶与突变分析
为了对PaNOMT9/4/SAM复合物进行结晶,纯化的PaNOMT9蛋白(20 mg/ml)与5 mM(S)-coclaurine(4)和5 mM SAM共同孵育。然后,通过静滴气相扩散法在4°C下生长晶体。沉淀剂井液由20% w/v PEG1000、100 mM硫酸锂一水合物和100 mM柠檬酸三钠二水合物(pH 5.5)组成。晶体逐渐转移到含有25%甘油的沉淀剂井液中,然后在液氮中快速冷冻以存储。所有数据集均在上海同步辐射光源(SSRF)BL18U1束流线的低温条件下(100 K)收集,结构通过分子置换法使用搜索模型6I70进行求解。然后使用Phenix软件进行模型优化(77),并在Coot软件中手动建模(78)。最终结构的优化结果显示,工作R因子(Rwork)为15.9%(自由R因子Rfree = 19.3%),几何学良好(见表S13)。
通过基因拼接重叠扩增PCR方法构建了PaNOMT9中选定残基的突变体,并使用底物4进行突变体活性测试(进行三次重复)。







:将域名邮箱添加至Outlook客户端)




![2025年渗透测试面试题总结-腾讯[实习]科恩实验室-安全工程师(题目+回答)](http://pic.xiahunao.cn/2025年渗透测试面试题总结-腾讯[实习]科恩实验室-安全工程师(题目+回答))




)

